NEWS LETTER
ニュースレター詳細
新製品情報: HIV-1 and HIV-2 gp140 Antigens
記事発行元:Native Antigen Company
HIV診断のための新しいツール
生命を脅かす後天性免疫不全症候群(AIDS)の蔓延を抑え、人々を守る唯一の手段は、依然として高感度で利用しやすい診断薬と薬物療法である。ヒト免疫不全ウイルス1(HIV-1)は世界的にAIDSの最も一般的な原因であり、HIV-2感染は主に西アフリカの人々に限られている。これらのHIV感染症の臨床的特徴は異なり、HIV-1に感染した患者は十分な治療と早期診断なしにAIDSに進行する可能性が高い(Nyamweya et al.) HIV-1とHIV-2のウイルス侵入受容体はエンベロープタンパク質gp160であり、成熟糖タンパク質gp120と膜貫通タンパク質gp41を特徴とする膜貫通型糖タンパク質である。gp160はポリタンパク質として合成され、宿主に常在するプロテアーゼfurinによって切断される(Checkley et al.) ビリオン内ドメインと膜貫通ドメインを除去すると、gp140と呼ばれる可溶性の分泌型gp160ができる。The Native Antigen Company (TNAC)は現在、タグなしHIV-1糖タンパク質gp140グループMサブタイプA、B、C、CRF01-AE、グループO、およびHIV-2糖タンパク質gp140を提供しており、新世代の診断アッセイと構造研究を可能にしている。すべてのタンパク質は、3つのアミノ酸の変化(一般にSOSIPと呼ばれる)を利用して修飾され、ジスルフィド橋がgp120とgp21と呼ばれるgp41の可溶性ドメインを共有結合し、複合体の安定性と寿命を向上させている(Binleyら、2000)。
分類と疫学
HIV-1の感染性は、このウイルスの遺伝的不均一性の広さに起因しており、その結果、様々な世界的分布を持つこのウイルスの多くの亜種が生成されている。HIV-1亜種の4つの主要な系統群には、メイングループ(M)、アウトライアーグループ(O)、非MまたはOグループ(N)、そして最近加わったグループPがある(Vallari et al.)世界中のHIV-1感染のほとんどは、A-K亜型に分類されるメイングループに関連している。O群、N群、P群はHIV-1感染者全体の約1%に過ぎないと推定されるが、M群の亜型はHIV-1感染者全体の約95%を占めている(Giovanetti et al.)
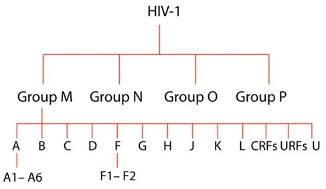
図1. HIV-1株間の進化的関係を図式化したもの。上から、4つの主要なグループ、サブタイプ(A-U)、サブサブタイプ(例えば、A1-A-6)があり、「U」は型付けされていないものを意味し、「CRFs」は循環組換え型(circulating Recombinant Forms)の略称、「URFs」はユニーク組換え型(Unique Recombinant Forms)を意味する。写真出典:Giovanetti et al: Giovanetti et al.
C亜型は最も流行しているウイルス型で、主にインドやエチオピアのような低所得国や南部アフリカの感染症例に関連しており、そこでは成人の4人に1人がHIV-1感染症に罹患している。ヨーロッパとアメリカ大陸で最も流行しているHIV-1サブタイプBに関連する宣伝や研究への関心は不釣り合いに高いが、HIV-1感染全体の約12%を占めるにすぎない(Hemelaar et al.)
CRF01_AE亜型と他のいくつかのCRFは、東アジアと東南アジアにおける主なHIV-1亜型である。この地域における様々なCRFの急速な出現は、異なるリスクグループにおけるB亜型とCRF01_AEの共流行の間に始まった。その結果、タイ、マレーシア、シンガポールで最初に見つかったCRF15_01B11やCRF34_01Bなど、これらの亜型からさらに複雑な組み換え株が出現した(Liu et al.)
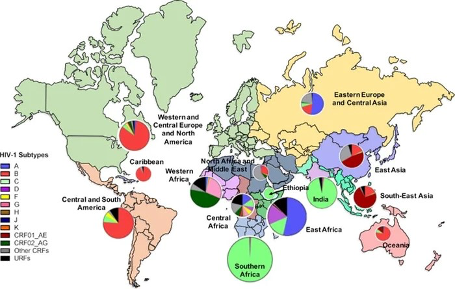
図2. 世界のHIV-1 M群亜型の有病率。写真出典: Gartnerら、2020年。
ウイルスの侵入
HIV-1
ウイルスエンベロープタンパク質gp160はHIV-1の侵入の中心であり、膜融合とそれに続く宿主細胞への遺伝子コアの送達を可能にする。HIVエンベロープタンパク質が宿主細胞膜に接触すると、宿主レセプターCD4およびコアセプターCCR5/CXCR4に結合する。この結合がエンベロープ糖タンパク質3量体の多段階構造変化を引き起こす。これらの構造変化によって融合ペプチドが露出し、その後のウイルスと宿主細胞膜の融合とゲノムの宿主細胞質への侵入が可能になる(Wang et al.)
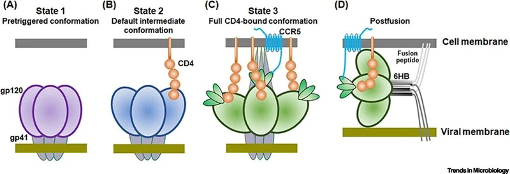
図3. ウイルス侵入に至るHIVエンベロープタンパク質のコンフォメーションの段階。写真出典:Wang et al: Wangら、2020年。
HIV-1エンベロープ糖タンパク質複合体の6つのサブユニット間の自然な非共有結合相互作用では、組換え発現タンパク質の三量体構造を維持することはできない。TNACの顧客は、組換えgp41抗原単独では一般的に不溶性であり、高濃度のイオン性洗浄剤を含む緩衝液なしでは取り扱いが困難であるため、体外診断用医薬品のアッセイ製造には不向きであると報告しています。
HIVアッセイ製造の課題に対する当社のソリューション
gp120とgp41の間にジスルフィド結合を導入することで、HIV-1糖タンパク質複合体のネイティブライクな構造を安定化させ、gp120とgp41可溶性ドメインであるgp21を橋渡しする。得られたgp140抗原(SOSIP)は、ネイティブなHIVエンベロープタンパク質の抗原特性を模倣しており、中和抗体応答の主要な標的として、アッセイ開発やワクチンデザイン研究において重要である(Pugach et al.) 組換えgp140タンパク質は可溶性で、単純な生理的緩衝液中で安定であるため、アッセイ法の開発や生産に便利である(Harris et al.) ネイティブなフリン切断部位のアミノ酸配列を変更することにより、gp140ポリタンパク質の切断を促進し、Binleyら、2000に記載されているように、gp120:gp21エクトドメイン抗原性標的の最適な提示をもたらした。

図4. 我々のgp140(SOSIP)は、タンパク質の成熟を可能にする強化されたフリン切断部位(Binley et al.、2000)と、gp120とgp21の間のジスルフィド結合を特徴とし、共有結合を維持する(Pugach et al.、2015)。
The Native Antigen Company (TNAC)は、以下のウイルスグループおよびサブタイプのHIV-1およびHIV-2 gp140(SOSIP)抗原の上市を発表いたします。
| Cat. No. | Product | Unit |
|---|---|---|
| REC32083 | Human immunodeficiency virus subtype A, gp140 (SOSIP), untagged | 100ug/500ug |
| REC32084 | Human immunodeficiency virus subtype C, gp140 (SOSIP), untagged | 100ug/500ug |
| REC32085 | Human immunodeficiency virus subtype AE, gp140 (SOSIP), untagged | 100ug/500ug |
| REC32086 | Human immunodeficiency virus subtype B, gp140 (SOSIP), untagged | 100ug/500ug |
| REC32076 | Human immunodeficiency virus type 2, gp140 (SOSIP), untagged | 100ug/500ug |
| REC32082 | Human Immunodeficiency virus clade O, gp140 (SOSIP), untagged | 100ug/500ug |
References
- Binley, J.M. et al. (2000) ‘A recombinant human immunodeficiency virus type 1 envelope glycoprotein complex stabilized by an intermolecular disulfide bond between the gp120 and GP41 subunits is an antigenic mimic of the trimeric virion-associated structure’, Journal of Virology, 74(2), pp. 627–643. doi:10.1128/jvi.74.2.627-643.2000.
- Checkley, M.A., Luttge, B.G. and Freed, E.O. (2011) ‘HIV-1 envelope glycoprotein biosynthesis, trafficking, and incorporation’, Journal of Molecular Biology, 410(4), pp. 582–608. doi:10.1016/j.jmb.2011.04.042.
- Gartner, M.J. et al. (2020) ‘Understanding the mechanisms driving the spread of subtype C HIV-1’, EBioMedicine, 53, p. 102682. doi:10.1016/j.ebiom.2020.102682.
- Giovanetti, M. et al. (2020) ‘Molecular epidemiology of HIV-1 in African countries: A comprehensive overview’, Pathogens, 9(12), p. 1072. doi:10.3390/pathogens9121072.
- Harris, A. et al. (2011) ‘Trimeric HIV-1 glycoprotein GP140 immunogens and native HIV-1 envelope glycoproteins display the same closed and open quaternary molecular architectures’, Proceedings of the National Academy of Sciences, 108(28), pp. 11440–11445. doi:10.1073/pnas.1101414108.
- Hemelaar, J. et al. (2019) ‘Global and regional molecular epidemiology of HIV-1, 1990–2015: A systematic review, global survey, and Trend Analysis’, The Lancet Infectious Diseases, 19(2), pp. 143–155. doi:10.1016/s1473-3099(18)30647-9.
- Liu, Y. et al. (2012) ‘Identification of a novel HIV type 1 circulating recombinant form (CRF52_01B) in Southeast Asia’, AIDS Research and Human Retroviruses, 28(10), pp. 1357–1361. doi:10.1089/aid.2011.0376.
- Marcelino, J.M. et al. (2006) ‘Use of a new dual-antigen enzyme-linked immunosorbent assay to detect and characterize the human antibody response to the human immunodeficiency virus type 2 envelope GP125 and GP36 glycoproteins’, Journal of Clinical Microbiology, 44(2), pp. 607–611. doi:10.1128/jcm.44.2.607-611.2006.
- Nyamweya, S. et al. (2013) ‘Comparing HIV-1 and HIV-2 infection: Lessons for viral immunopathogenesis’, Reviews in Medical Virology, 23(4), pp. 221–240. doi:10.1002/rmv.1739.
- Pugach, P. et al. (2015) ‘A native-like sosip.664 trimer based on an HIV-1 subtype B env gene’, Journal of Virology, 89(6), pp. 3380–3395. doi:10.1128/jvi.03473-14.
- Sanders, R.W. et al. (2013) ‘A next-generation cleaved, soluble HIV-1 env trimer, BG505 SOSIP.664 gp140, expresses multiple epitopes for broadly neutralizing but not non-neutralizing antibodies’, PLoS Pathogens, 9(9). doi:10.1371/journal.ppat.1003618.
- Valadés-Alcaraz, A., Reinosa, R. and Holguín, Á. (2022) ‘HIV transmembrane glycoprotein conserved domains and genetic markers across HIV-1 and HIV-2 variants’, Frontiers in Microbiology, 13. doi:10.3389/fmicb.2022.855232.
- Vallari, A. et al. (2011) ‘Confirmation of putative HIV-1 Group P in Cameroon’, Journal of Virology, 85(3), pp. 1403–1407. doi:10.1128/jvi.02005-10.
- Wang, Q., Finzi, A. and Sodroski, J. (2020) ‘The conformational states of the HIV-1 envelope glycoproteins’, Trends in Microbiology, 28(8), pp. 655–667. doi:10.1016/j.tim.2020.03.007.

FAX:06-6152-9941